戞擇惗壔儅僯儏傾儖栚師
侾.偼偠傔偵
丂抈敀幙偺惛惢偼惗壔妛偺嵟傕婎慴揑側庤媄偱偁傝丄偙傟偵廗弉偡傞偙偲偼懠偺媄弍傪廗摼偡傞忋偱傕栶棫偮丅堚揱巕岺妛偺婎慴傕峺慺斀墳偱偁傝丄傑偨妀撪偁傞偄偼嵶朎幙撪偺彅抈敀偑揮幨傗東栿傪嵟廔揑偵偼惂屼偟偰偄傞丅屄乆偺惛惢曽朄偼徻偟偔弎傋側偄偑丄抈敀惛惢乮峺慺惛惢乯偵摉偨偭偰偼桳柤側僐乕儞僶乕僌攷巑偺10儠忦傪偙偙偵徯夘偡傞乮弌揟丗忋揷崙姲攷巑丄僗僞儞僼僅乕僪戝妛乯丅
Commandments for the enzymologists by Arthur Kornberg
1. Enzyme purification is its own reward.
2. The sabbath is a working day.
3. Ammonium sulfate is kind to enzymes.
4. The purification table is as important as the purification.
5. Dependency of pH comes before Ph.D.
6. You can't purify enzymes with toothpicks.
7. The cell is irrelevant to the isolated enzymes.
8. Don't waste clean thoughts on dirty enzymes.
9. Save the side fractions for handouts.
10. To spare the pen is to spoil the data.
俀.抈敀惛惢傪巒傔傞慜偵
俙丏栚揑
丂抈敀偺惛惢偵摉偨偭偰偼栚揑傪偼偭偒傝偝偣傞偙偲偑戝愗偱偁傞丅晹暘惛惢偱傛偄偺偐丄姰慡偵扨堦側傕偺偑昁梫偐丄峈懱嶌惉偵巊偆偺偐丄儁僾僠僪攝楍傪抦傞昁梫偑偁傞偺偐丄摍偱偁傞丅惛惢偦偺傕偺偱榑暥偵偡傞偲偒偼昁偢怴偟偄峺慺偱側偔偰偼側傜側偄丅庬嵎偺巇帠偼堦棳偺崙嵺帍偵偼嵦戰偝傟側偄丅
俛丏憻婍傗嵶朎偺慖戰
丂栚揑偵墳偠偰憻婍傗嵶朎傪慖戰偡傞丅弌敪帪偺斾妶惈乮栚揑偲偡傞抈敀傪慡抈敀偱妱偭偨傕偺乯偺堘偄偑丄偦偺屻偺峴掱偺嬯楯傪寛掕偡傞丅
俠丏埨掕惈偺専摙
丂峺慺傪弶傔偲偡傞抈敀偼曄惉偟丄惗暔妶惈傪幐偆丅廬偭偰丄偳偺傛偆側忦審偱抈敀偑嵟傕埨掕偐傪挷傋傞偙偲偑娞怱偱偁傞丅捠忢丄彮検偺僒儞僾儖傪梡偄偰丄幒壏丄係搙C丄椻搥摍傪峴偄埨掕搙傪僠僃僢僋偡傞丅傑偨丄埨掕壔嵻傪尒偮偗傞偙偲抈敀惛惢惉岟偺尞偱偁傞丅
俢丏妶惈應掕曽朄偺奐敪
丂抈敀惛惢偵摉偨偭偰偼壗傜偐偺巜昗偑昁梫偱偁傞丅峺慺偺応崌丄偦傟偑峺慺妶惈應掕朄偺奐敪偱偁傞丅應掕偼惓妋偱側偔偰偼側傜側偄偑丄傛傝廳梫側偙偲偼娙扨偱恦懍偵偱偒傞曽朄偱側偔偰偼側傜側偄丅側偤側傜丄妶惈應掕偵帪娫傪梫偡傞偙偲偼偦偺傑傑抈敀偺幐妶傪堄枴偡傞偐傜偱偁傞丅
俤丏奅柺妶惈暔幙乮Detergent乯偺慖戰
丂枌峺慺偺応崌偼偳偺傛偆側奅柺妶惈壔嵻傪梡偄傞偐偑嵟傕娞怱偱偁傞丅奅柺妶惈壔嵻偵偼暿昞偺條側傕偺偑偁傞丅帀梟惈丄悈梟惈偺僶儔儞僗傪帵偡傕偺偲偟偰HLB僫儞僶乕乮hydrophilicity
lipophilicity balance number)偑懚嵼偟丄壜梟壔嵻慖戰偺偲偒偺堦偮偺婎弨偲側傞丅枌抈敀偼壜梟壔偝傟側偄偲惛惢偱偒側偄丅壜梟壔偺巜昗偼堦斒偵偼師偺嶰偮偺僋儔僀僥儕傾偑偁傞丅戞堦偵侾侽枩g偱墦怱偟偰忋惔偵偔傞偙偲丅戞擇偵儃傾僒僀僘偺戝偒側僎儖傠夁僇儔儉偱拞偵偼偄傞偙偲乮void
volume偵偱側偄偙偲乯丄偦偟偰戞嶰偵0.45mm僒僀僘偺僼傿儖僞乕傪捠傞偙偲偑忋偘傜傟傞丅妶惈壔嵻偼扨撈偱丄偁傞偄偼慻傒崌傢偝傟偰巊梡偝傟丄堦斒偵惛惢偑恑傓偲偲傕偵擹搙傪敄偔偡傞丅壜梟壔嵻傪梡偄偰丄枌偐傜棧偡偲摨帪偵抈敀偼晄埨掕偲側傞丅偟偨偑偭偰丄奅柺妶惈壔嵻偲埨掕壔嵻偺椙偒慻傒崌傢偣偺敪尒偑惛惢惉岟偺戝慜採偱偁傞丅
俁..抈敀惛惢偺尨棟
丂抈敀幙偼傾儈僲巁偺廤崌懱偱偁傝丄偙傟偵摐嵔傗帀幙偑嫟桳寢崌偟偨傕偺偱偁傞丅廬偭偰丄偙傟傪暘棧偡傞偺偼抈敀偺帩偮丄摍揹揰丄僒僀僘丄梟夝搙乮悈丄桳婡梟攠拞側偳乯丄摿暿側暔幙傊偺恊榓惈乮峺慺偺応崌丄婎幙丄曗峺慺側偳乯摍傪棙梡偡傞丅徻嵶側曽朄偼徣棯偡傞偑丄偙偙偱偼堦斒揑尨棟偲拲堄揰傪婰嵹偡傞丅
俙丏梟夝搙偵傛傞暘棧
丂戙昞揑側傕偺偵棸巁傾儞儌僯僂儉傪梡偄偨傕偺偑偁傝丄棸埨乮偲棯偡乯暘巕偵悈暘巕偑偲傜傟偨寢壥丄抈敀偺梟夝惈偑棊偪傞惈幙傪棙梡偟偰偄傞丅堦斒偵偼慳悈惈偺傕偺傎偳捑揳偟傗偡偔丄傑偨暘巕検偺戝偒偄傕偺傎偳捑揳偟傗偡偄丅摨偠暘巕検偺傕偺偱傕梟夝搙偵嵎偑偁傞偨傔丄偙偺曽朄偑梡偄傜傟傞丅偙偺曽朄偼抈敀傪捑揳偟偨偁偲丄擟堄偺梕検偵梟偐偡偙偲偑偱偒傞偨傔丄擹弅偵曋棙偱偁傝丄偝傜偵梟塼傪師偺僇儔儉偵崌傢偣偨娚徴塼偱摟愅偡傞偨傔丄惛惢偺戞堦抜奒偵曋棙偱偁傞丅弶傔俀侽丄係侽丄俇侽亾...偲偄偆傛偆偵戝傑偐偵暘夋偟丄偦傟偧傟偺暘夋偺妶惈偲抈敀傪應掕偟偨屻丄嵟廔揑側暘夋偺斖埻傪寛傔傞丅
丂摨偠傛偆側尨棟偱梟夝搙偺嵎傪棙梡偟偨傕偺偵桳婡梟攠偵傛傞捑揳暘棧偑偁傞丅堦斒偵偼傾僙僩儞偐僄僞僲乕儖偑梡偄傜傟傞丅偙偺挿強偼帀幙側偳偺崿擖傪彍偔偙偲偑偱偒傞偙偲偱偁傝丄傑偨丄摟愅傕晄梫偱偁傞偙偲偱偁傞乮拏慺僈僗偺悂偒晅偗乯丅棸埨暘夋偑偆傑偔峴偐側偄偲偒偵師偵帋偝傟傞丅抁強偼抈敀偑曄惉乮旕壜媡揑乯偟傗偡偄偙偲偱偁傝丄梟塼丄梕婍丄墦怱婡偼偄偢傟傕侽並C埲壓偵曐偮昁梫偑偁傞丅僄僞僲乕儖丄傾僙僩儞側偳偺桳婡梟攠偲棸埨偺亾偼堄枴偑偪偑偄丄桳婡梟攠偺応崌偼vol/vol偱偁傝丄棸埨偼朞榓棸埨梟塼傪侾侽侽亾偲偟偨偲偒偺憡懳抣偱偁傞丅棸埨偺亾偼昞侾傪嶲徠偝傟偨偄丅
俛丏僀僆儞岎姺僋儘儅僩僌儔僼傿乕偲摍揹揰僋儘儅僩僌儔僼傿乕
丂抈敀偼傾儈僲巁傗摐偺懡暘巕懱偱偁傞丅屄乆偺傾儈僲巁傗摐幙偺堦晹偼亄偁傞偄偼-偺壸揹傪桳偟偰偄傞乮傾儈僲婎丄偁傞偄偼僇儖儃僉僔儖婎乯丅偙傟傜傪憤寁偟偨傕偺傪偦偺抈敀偺摍揹揰乮pI,
isoelectric point)偲尵偄丄偦偺pH偱抈敀偼揹婥揑偵拞惈偲側傞丅懡偔偺抈敀偼巁惈壸揹偱偁傝丄pI偼俆埲壓偱偁傞丅偄傑丄pI偑俆偺抈敀傪pH俉偺娚徴塼偵梟偐偡偲丄抈敀偼堿僀僆儞偲側傞丅偙偺堿僀僆儞傪媧拝偡傞庽帀傪堿僀僆儞岎姺(anion
exchanger)庽帀偲徧偟丄堦斒偵傾儈僲婎傪桳偟偰偄傞乮椺:DEAE, QAE, MonoQ)丅媡偵僇儖儃僉僔儖婎乮CM)傗棸巁婎(SM,
MonoS)傪帩偮傕偺傪梲僀僆儞岎姺庽帀偲屇傇丅僇儔儉僋儘儅僩僌儔僢僢僼傿乕傪峴偆偵摉偨偭偰偼傑偢丄庽帀偵栚揑偲偡傞抈敀偑寢崌偡傞偐偳偆偐傪僠僃僢僋偟丄偮偄偱丄暘棧偺曽朄傪峫偊傞丅堦斒偵暘棧偵偼墫(NaCl)偺擹搙岡攝傗pH偺曄壔摍傪梌偊傞丅僀僆儞岎姺僋儘儅僩僌儔僼傿乕偑昞柺壸揹偵傛傝斀墳偡傞偺偵懳偟偰丄摍揹揰偦偺傕偺偱暘偗傞僇儔儉偑偁傝丄嫵幒偵偼Mono亅P僇儔儉屇偽傟傞傕偺偑偁傞丅
俠丏僎儖傠夁僋儘儅僩僌儔僼傿乕
丂惓妋偵偼暘巕僒僀僘乮暘巕検偱偼側偔乯偺堘偄傪棙梡偡傞暘棧曽朄偱偁傞丅偄偢傟傕栐栚忬丄偁傞偄偼棻巕偺拞偵寠(pore)偑嬻偄偰偍傝丄僒僀僘偺戝偒側傕偺偼偦偺奜傪捠夁偡傞偨傔憗偔堏摦偟丄彫偝側傕偺偺傒偑丄拞傪捠傝丄堏摦懍搙偑抶偔側傞偙偲傪棙梡偟偰偄傞丅僎儖偺扲懱偵傛傝僙僼傽僨僢僋僗丄僙僼傽僋儕儖丄僗乕僷乕儘乕僗摍偑偁傝丄傑偨暘棧偺斖埻傪寛傔傞暘巕僒僀僘偱偄偔偮傕偺庬椶偑偁傞丅僙僼傽僨僢僋僗G俀侽侽偼惷悈埑乮娚徴塼偺儕僓乕僶乕偺埵抲偲僇儔儉偺傾僂僩儗僢僩偺埵抲偺嵎乯偑侾侽cm埲忋偵側傞偲栚媗傑傝偑婲偙傝丄媫懍偵棳懍偑棊偪傞偺偱拲堄偡傞丅堦斒偵僎儖偺扲懱偲偺僀僆儞惈偺憡屳嶌梡傪尭傜偡偨傔丄0.1M掱搙偺NaCl傪壛偊傞偙偲偑懡偄丅
俢丏慳悈惈僋儘儅僩僌儔僼傿乕
丂抈敀偵偼悈傊偺梟偗傗偡偝偺巜昗偲偟偰慳悈惈偑偁傞丅堦斒偵巁惈偁傞偄偼墫婎惈偺傾儈僲巁丄偦偟偰拞惈傾儈僲巁偺偆偪懁嵔偺僸僪儘僇乕儃儞偺抁偄傕偺乮傾儔僯儞丄僌儕僔儞乯摍偼怴悈惈偱偁傝丄媡偵儘僀僔儞丄償傽儕儞丄僀僜儘僀僔儞側偳懁嵔偺挿偄傕偺偼慳悈惈偱偁傞丅偙傟傜傾儈僲巁偺憤悢偑抈敀偺慳悈惈傪寛掕偡傞偲峫偊傜傟傞偑丄偙偺堘偄傪棙梡偟偨傕偺偑慳悈惈僇儔儉偱偁傝丄庽帀偵慳悈婎偑寢崌偟偰偄傞丅寢崌椡偼僀僆儞擹搙偑崅偄傎偳乮僀僆儞寢崌偑梷偊傜傟偰乯崅偔側傞偺偱丄墫傗棸埨傪壛偊偰丄寢崌偝偣丄偙傟傜傪尭傜偡岡攝傪偐偗偰梟弌偡傞丅嫵幒偵偁傞僇儔儉偱偼僆僋僠儖乮C偑俉乯亜僼僃僯儖乮C偑俇乯偺弴偱寢崌偑嫮偄丅偝傜偵梟弌偵摉偨偭偰偼僄僠儗儞僌儕僐乕儖丄傾儖僐乕儖丄奅柺妶惈壔嵻摍傪壛偊偰梟弌偟傗偡偔偡傞偙偲傕偁傞丅
俢丏媧拝僇儔儉僋儘儅僩僌儔僼傿乕
丂僴僀僪儘僉僔傾僷僞僀僩偵戙昞偝傟傞傕偺偱丄僇儖僔僂儉偲儕儞巁傊偺僀僆儞寢崌搙偺憤榓傪棙梡偟偰偄傞丅僸僪儘僉僔傾僷僞僀僩偼Ca10(PO4)6(OH)俀偺寢徎偱偁傞丅僀僆儞岎姺僇儔儉偲摨偠埖偄偱偁傞偑丄晛捠偼侾乕俆mM偺儕儞巁娚徴塼(pH6.8)偱寢崌偝偣丄400mM傑偱偺摨娚徴塼偱傎偲傫偳偡傋偰偺抈敀偑梟弌偝傟傞丅惛惢偺屻傠偺抜奒偱巊梡偝傟傞丅傑偨丄巁偵偼庛偄偺偱pH傪俆埲壓偵偟偰偼偄偗側偄丅乮愻忩帪傕巁傪梡偄偰偼偄偗側偄乯丅
俤丏傾僼傿僯僥傿乕僋儘儅僩僌儔僼傿乕
丂偦偺抈敀傗峺慺偵摿堎揑偵寢崌偡傞暔幙傪僇儔儉偵偮偗丄偙傟偵傛傝栚揑偲偡傞埲奜偺抈敀傪偡傋偰慺捠傝偝偣傛偆偲偡傞帋傒偱偁傞丅偆傑偔峴偗偽侾僗僥僢僾偱栚揑暔傪偲傞偙偲傕偱偒傞丅峺慺偺応崌丄婎幙丄曗峺慺傪寢崌偝偣偨傝丄偁傞偄偼抈敀偺扨僋儘乕儞峈懱傪棙梡偟偨傝丄傑偨庴梕懱偺応崌儕僈儞僪傪偮偗傞側偳偺曽朄偑偁傞丅偙傟傜帺嶌偺僇儔儉偺傎偐丄嫵幒偵偁傞傕偺偼僽儖乕僙僼傽儘乕僗(NAD,NADP椶帡懱乯丄僿僷儕儞僙僼傽儘乕僗乮惉挿場巕丄儂僗儂儕僷乕僛側偳偺惛惢偵梡偄傞乯丄僌儖僞僠僆儞僙僼傽儘乕僗(GST偲偺梈崌抈敀偺惛惢乯側偳偑偁傞丅傑偨丄僒僂僗僂僄僗僞儞朄傗俢俶俙寢崌場巕偺惛惢摍傕栚揑偲偡傞僰僋儗僆僠僪傊偺媧拝傪棙梡偟偨傕偺偱偁傞丅
俥丏媡憡僇儔儉僋儘儅僩僌儔僼傿乕
丂扲懱偲偟偰C係丄C俉丄C侾俉乮俷DS乯摍偑偁傝丄偙傟傪悈偲傾僙僩僯僩儕儖偺岡攝偱梟弌偡傞丅堦庬偺慳悈惈僇儔儉偱偁傞丅寢崌椡偼扽慺悢偑戝偒偄傎偳嫮偔丄廬偭偰崅擹搙偺傾僙僩僯僩儕儖傪昁梫偲偡傞丅抈敀偺壸揹傪側偔偟丄僀僆儞寢崌偵傛傞姳徛傪攔彍偡傞栚揑偱捠忢0.1亾偺俿俥俙(trifluoroacetic
acid)偑壛偊傜傟傞丅傾僙僩僯僩儕儖偺戙傢傝偵僀僜僾儘僺儖傾儖僐乕儖傕巊傢傟傞丅媡憡僇儔儉偼僔儕僇僎儖側偺偱丄傾儖僇儕(pH>10)偵偟偰偼側傜側偄丅
係.偦偺懠偺彅拲堄
丂嵶朎幙峺慺偼堦斒偵偼棸埨暘夋乮妀偑懡偄偲巚傢傟傞傕偺丄椺偊偽嵶朎摍偼偙偺慜偵僗僩儗僾僩儅僀僔儞張棟偱彍妀巁傪峴偆偲傛偄乯仺摟愅仺僀僆儞岎姺僇儔儉仺僎儖傠夁僇儔儉仺媧拝偁傞偄偼傾僼傿僯僥傿乕僇儔儉摍偺弴斣偱恑傔傞偙偲偑懡偄丅摨偠僇儔儉傪擇搙巊偆偺傕桳岠偱偁傞丅
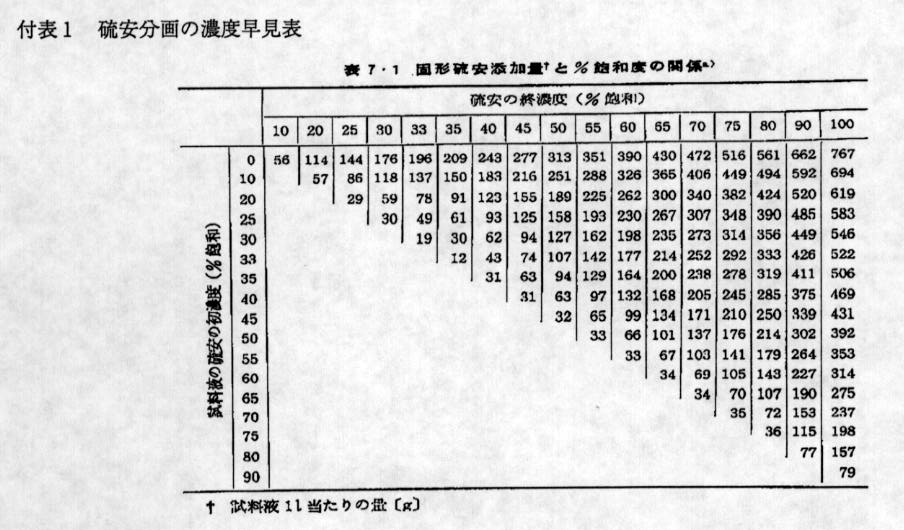
晅昞侾丂棸埨暘夋偺擹搙憗尒昞(JPEG 128K)
晅昞俀丂戞擇惗壔偱強桳偡傞僇儔儉堦棗昞(偐偭偙撪偺悢帤偼捈宎x崅偝丄扨埵mm乯
堿僀僆儞岎姺僇儔儉
MonoQ(5x50), MonoQ(10x100), TSKDEAE-5PW(7.5x75,丂8x75丄掇捉そ幂诮),
Polyanion SI丂(5x50),(10x100)
Q-Sepharose(open column)
乮拲堄乯堦斒偵壜梟惈抈敀偵偼MonoQ椙偄偑丄媧拝偑懡偄丅Polyanion偼暘棧偑僔儍乕僾偱偁傞丅偟偐偟丄僔儕僇僎儖惈偺偨傔傾儖僇儕偱偺愻忩偑偱偒偢丄晄曋偱尰嵼偼敪攧偝傟偰偄側偄丅MonoQ偺戝偒偄傕偺偼敪攧摉弶侾侽侽枩墌丄尰嵼偱傕係侽枩墌偡傞崅壙側傕偺側偺偱丄庢埖偵拲堄丅
梲僀僆儞岎姺僇儔儉
Mono S(5x50), MonoS(10x100), TSKSP-5PW(7x50)
乮拲堄乯忋偲摨偠
僎儖傠夁僇儔儉
Superose 12(10x300), Superose 12(30x500),TSKG3000XL(7.8x300),
TSKG3000SW(7.5x300,7.5x600)
Superdex
扲懱偲偟偰偼Sephadex G-25, Sephadex G-50, Sephacryl S-300, Sephacryl
S-1000
乮拲乯僎儖傠夁偼暘巕検偵傛傝條乆側庬椶偑偁傞偺偱丄栚揑偲偡傞抈敀偺暘巕検偑梊憐偝傟偨傜丄偦傟偵崌偆傕偺傪巊梡偡傞丅Sephacryl偼懠偺傕偺偲斾傋偰傗傗媧拝偑懡偄丅
慳悈惈僇儔儉丂丂
Phenyl-superose(5x50), TSK Ether 5PW(7.5x75), TSK Phenyl-5PW(7.5x75),
僸僪儘僉僔傾僷僞僀僩僇儔儉
TSK HA-1000(7.5x75)
乮拲乯巁惈偵偟偰偼偄偗側偄丅(pH偼俆埲忋偱巊偆乯
媡憡僇儔儉
PEP-PRC, TSK-ODS(C18)奺庬
乮拲乯暘棧偼椙岲偩偑丄桳婡梟攠傪巊偆偨傔堦斒偵妶惈偼幐偆丅
傾僼傿僯僥傿乕僇儔儉
TSK-Blue-5PW(7.5x750), Heparin-sepharose(7.5x750)
晅昞俁丂奅柺妶惈嵻偺庬椶偲HLB僫儞僶乕(JPEG64K)
晅昞4丂奅柺妶惈嵻偺庬椶偲HLB僫儞僶乕(JPEG96K)
晅昞5丂奅柺妶惈嵻偺庬椶偲HLB僫儞僶乕(JPEG96K)
戞擇惗壔儅僯儏傾儖栚師
{kind=link}
{kind=link}
{kind=link}
{kind=link}